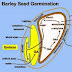
Plant Physiology Role of ABA and Components of the ABA Signalling Pathway Membangun Indonesia 5:59 PM